
刊行物





- HOME
- 刊行物のご案内
- 心理学ワールド
- 108号 行動主義を見つめなおす――心なき心理学と呼ばれて
- プラナリアの学習と記憶の研究─マコーネルのエピソードから学んだこと
【小特集】
プラナリアの学習と記憶の研究─マコーネルのエピソードから学んだこと
田積 徹(たづみ とおる )
Profile─田積 徹
同志社大学大学院文学研究科博士後期課程修了。博士(心理学),博士(医学)。2012年より現職。専門は行動学的神経科学,実験心理学。著書に『私たちの心理学』(共編,川島書店),『情動の仕組みとその異常』(分担執筆,朝倉書店)など。
プラナリアとは?
プラナリアという用語は,扁形動物門のウズムシ綱(類)において区別されている13目のうち,中・大型の動物群を指す総称である[1]。図1は筆者が研究室で維持・飼育している3種のプラナリアである。プラナリアは自切による無性生殖や卵殻(コクーン)を産む有性生殖によって増えていく。図1のリュウキュウナミウズムシは生殖器官を持っていないが,有性個体をエサとして与えることで人為的に有性化ができる。

プラナリアの研究史
プラナリアの行動学的研究はヴァン・オーイェ[2]によって始められたといわれている[3]。その研究では,水中を浮遊しないプラナリアの特性を利用した迷路が開発されている。
その後,1950年代から1960年代にわたり,世間の注目を浴びたプラナリアの学習と記憶の研究がマコーネルらによって報告された。彼らは光を条件刺激,電気ショックを無条件刺激として対提示すると,光の提示に対する条件反応をプラナリアが獲得することを報告した[4]。ほかにも,彼らは条件反応を獲得したプラナリアを頭部と尾部に切断し,尾部から頭部を再生した個体も条件反応が保持されていること[5]や,学習した個体を共食い[6],あるいは,学習した個体から抽出されたRNAを注射[7]した実験経験のないプラナリアも学習を保持することを報告した。このような現象はRNA媒介記憶転移と呼ばれている。マコーネルはこれらの知見を喧伝し,それによって生じたその当時の様子が社会学者によって2006年に報告されている[8]。その報告によると,当時多くの人々がミミズやプラナリアなどのワームが学習できること,ましてや共食いやRNA注射を通じて記憶が転移できるとは信じておらず,さらにプラナリアでの記憶の転移の再現性が得られていない状況下であった。その中でマコーネルはメディアを通して,記憶が転移できる錠剤(記憶ピル)や注射が開発される未来について大々的に語った。さらに1960年代には,誰もが学習することを疑っていないラットにおいてRNA媒介記憶転移が生じることが複数のラボから報告された[9](当時,これらの報告に対しても再現性について疑念が示された[10])。ヒトに近いラットでこの記憶転移が示されたため,マコーネルの研究は見向きされなくなり,プラナリアの学習能力の信頼性までもが失われてしまった。
マコーネルの研究の信憑性
マコーネル以外の研究者も,プラナリアが古典的条件づけの獲得や阻止,隠蔽を示すことを報告している[11]。筆者も円形シャーレに少量の水とプラナリアを入れて,シャーレの円弧の側面を下にして,実験者が手でそのシャーレを傾けてプラナリアを水滴から遠ざけ,プラナリアがそこから水滴にたどり着くオペラント行動をシェイピングによって維持できることを確認している[12](図2)。

また,2001年には,RNA干渉がRNA媒介記憶転移のメカニズムである可能性が指摘された[13]。RNA干渉は細胞内で二本鎖RNAが少量コピーされるだけで生じるが,二本鎖RNAは細胞内での寿命が短く,RNA干渉を引き起こすには最小限の配列の長さが必要である。RNAは抽出のタイミングによって特徴が変動するので,マコーネルの研究で使用されたRNAがRNA干渉を誘発する上記の特徴を持っており,他のラボはそのような特徴を持っていないRNAを使用していたため再現できなかった可能性が指摘された。さらに,2018年に,長期的に鋭敏化したアメフラシのRNAを未訓練の個体に注入すると,その個体でも刺激に対する反射が鋭敏化することが報告された[14](図3)。
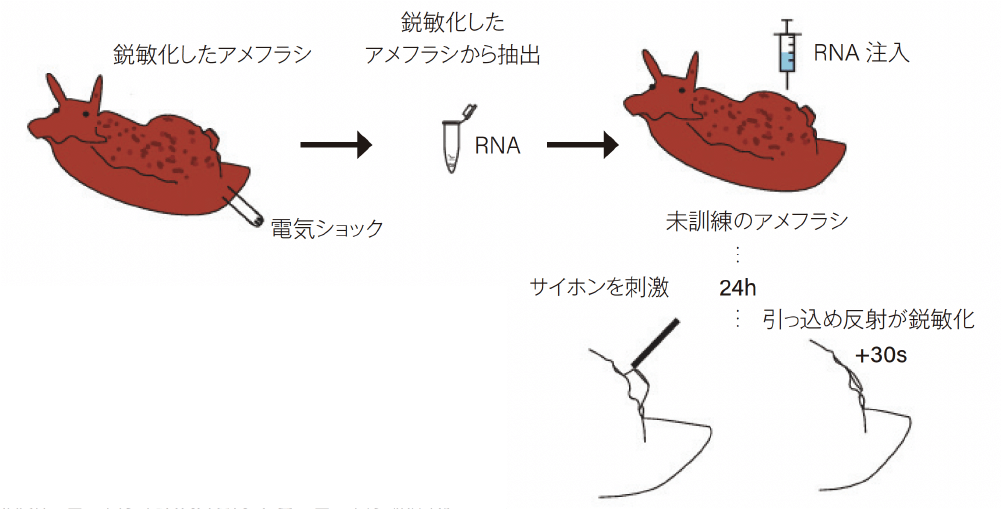
(文献[14]の責任著者の許可を得て図を一部複製)
マコーネルのエピソードから学ぶべきこと
上述したように,マコーネルが報告したプラナリアの学習は繰り返し再現性が確認され,再現性の疑われたプラナリアにおけるRNA媒介記憶転移についても,単に実験で使用されたRNAの特徴の違いに帰せられる可能性も指摘されている。にもかかわらず,マコーネルのエピソードは,RNA媒介記憶転移を再現できなかったという点が注目され,研究における戒めのニュアンスが込められた心理学の伝承(folklore)の一つとなっている[15]。このような捉えられ方をされるのは,マコーネルがメディアにおいて「記憶ピルが開発される未来」といった大衆受けするような夢物語を大々的に語ったことが理由の一つかもしれない。
昨年,スペインの学会に参加したときに聞いた発表では,無脊椎動物のカタツムリが痕跡条件づけを獲得したというデータが示され,その考察として,刺激の消失後に,その刺激の表象(イメージ)とその後に提示された別の刺激の間に連合ができる可能性があるという[16]。近年,無脊椎動物の認知に関する総説が発表され[17],無脊椎動物の認知能力や意識の有無を判断するための満たすべき条件が議論されている[18]。カタツムリがイメージを持つことができるのであれば,痕跡条件づけの獲得とは密接な関係があるだろう。しかしながら,カタツムリが痕跡条件づけを獲得できたのは,刺激の消失後も神経系の興奮状態が続いたために,その後に提示された刺激との連合ができたという別の説明が可能である。表象といった心的過程によって,その行動,とくに,動物の行動を説明するためには,そのような心的過程が介在しないとその行動は生じえないという行動課題の開発が必須であると筆者は考えている。そのような課題でもないのに,安易にその行動を心的過程で説明するのはマコーネルと同じ轍を踏んでいるのと変わらない。たしかに,カタツムリには表象する能力などないということを証明することは悪魔の証明であり,実質的に不可能であろう。しかし,それを証明することはできないからといって,別の説明が可能であるのに,カタツムリにはそのような能力があることを前提に,安易に心的過程で説明することは避けるべきである。プラナリアを利口な馬ハンスのようにしないためにも,これは大切なことだと思う。
文献
- 1.川勝正治他 (2007) 陸水学雑誌, 68, 461–469.
- 2.Oye, P. van. (1920) Natuurw Tijdschr, 2, 1.
- 3.Crawford, T. (1967) Behavioral modification of planarians. In W. C. Corning & S. C. Ratner (Eds.), Chemistry of learning: Invertebrate research (pp.234-250). Plenum Press.
- 4.Thompson, R., & McConnell, J. (1955) J Comp Physiol Psychol, 48, 65–68.
- 5. McConnell, J. V. et al. (1959) J Comp Physiol Psychol, 52, 1–5.
- 6.McConnell, J. V. (1962) J Neuropsychiat, 3(suppl 1), 542–548.
- 7.Zelman, A. et al. (1963) Worm Runner's Digest, 5, 14–21.
- 8. Stern, L. (2006) James V. McConnell’s advocacy of memory-transfer: Juggling three different personae–while walking a tightrope–without a net. Paper presented at the Thirty-Eighth Annual Meeting of Cheiron (The International Society for the History of Behavioral and Social Sciences).
- 9.Babich, F. R. et al. (1965) Science, 149, 656–657.
- 10.Byrne, W. L. et al. (1966) Science, 153, 658–659.
- 11.Prados, J. et al. (2013) Anim Cogn, 16, 177–186.
- 12.田積徹他 (2022) 人間科学研究, 44, 103–114.
- 13.Smalheiser, N. R. et al. (2001) Trends Neurosci, 24, 216–218.
- 14.Bédécarrats, A. et al. (2018) eNeuro, 5(3) ENEURO.0038–18.
- 15.Stern, L. (2010) Monitor on Psychol, 41, 28.
- 16.Muñiz-Moreno, J., & Loy, I. (2023) XXXIII International Conference of the Spanish Society for Comparative Psychology, Abstract Book, 33.
- 17.Menzel, R. et al. (2007) Cognition in invertebrates. In J. H. Kaas (Ed.), Evolution of nervous systems, vol II: Evolution of nervous systems in invertebrates (pp.403–422). Academic.
- 18.Jablonka, E. (2024) XXXIV International Conference of the Spanish Society for Comparative Psychology, Abstract Book, 16.
- *COI:本稿に関連して開示すべき利益相反はない。
PDFをダウンロード
1