
刊行物





【特集】
人を区別する
トラとライオン,人とロボット,自分の物と他人の物,……。あるものを他から区別することは生活に欠かせない活動です。そのなかでも私たちが得意とするのは,「人」の区別かもしれません。社会で生きるために,人は「人」を区別するメカニズムをもっています。このメカニズムは,目や耳に一瞬だけ入った情報からでも,それがどのような人かについての処理を促し,人種や性別,さらには人柄や能力などまで推測させようとします。ただし,これは意識して丁寧に行われる処理ではないため,エラーやバイアスをまねくこともあれば,差別,人命の軽視,社会の分断といった問題を生み出す要素にもなります。
本特集で取り上げる「人」の区別は,環境にある情報から人の動きを抽出する,乳幼児期から人の顔を見て様々な区別をする,自分と他者を極めて自然に区別する,仲間とそうでない者に異なる判断や処遇をするというものです。「人」を区別するメカニズムはどのようなものであり,私たちの生活にどのような意味や影響をもたらすのでしょうか。その謎に迫る研究をご覧ください。(大江朋子)
人の動きを区別する

平井真洋(ひらい まさひろ)
Profile─平井真洋
2005年,東京大学大学院博士課程修了。博士(学術)。東京大学特任研究員,日本学術振興会特別研究員,日本学術振興会海外特別研究員,愛知県心身障害者コロニー発達障害研究所研究員,自治医科大学医学部准教授などを経て,2020年より現職。専門は発達認知神経科学,実験心理学,身体化認知科学。著書は『社会的認知の発達科学(発達科学ハンドブック9)』(分担執筆,新曜社),『ソーシャルブレインズ』(分担執筆,東京大学出版会)など。
私たちは日常生活において,意識する・しないにかかわらず,他者の身体の動きに関する情報を解読している。例えば,警察の日常の活動を追跡したTV番組をご覧になったことがある読者の皆様は少なくないであろう。番組では,熟練した警察官が暗闇で「不審な挙動」をする人物を発見し,見事逮捕に至る話はよくある。この状況では,顔はおろか,身体の外見的な特徴も殆どわからないはずであり,まさに「動き情報」が不審者を特定する手がかりとなっている。このように,私たちは「他人の動き」に敏感である。
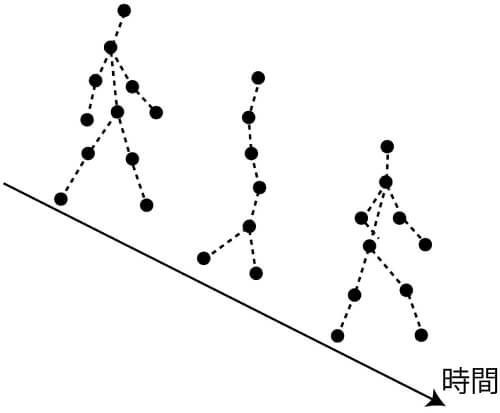
心理学を学んだことのある読者なら,スウェーデンの心理学者ヨハンソンによって報告されたBiological Motion(BM)と呼ばれる知覚現象(図1)はご存知であろう(Johansson, 1973)。BMは,ヒトの各関節に装着したわずか十数個の光点運動のみからヒトに関する様々な情報を解読することができる。例えば,光点運動のみから性別,意図,自己か他人か,騙そうとする動作,実際に重い荷物を持っているかどうかなどを判別できることがこれまで報告されている。
この知覚現象の発見から50年近く経つにもかかわらず,未だ多くの心理学者を魅了し,BMに関する研究論文が多数出版されている。2000年代に入り,BMを視覚刺激として用いた多くの脳機能イメージング研究が行われ,BM知覚処理を担う脳部位の空間的な同定が精力的に進められてきた。その結果,「社会的知覚」を担う社会脳の一部が関与することが明らかにされてきた。この発見から,後述するように一部の研究ではBMを社会的知覚処理に関するリトマス試験紙的な役割として用いるようにもなった。
このような動きに基づく一見すると複雑と思われる知覚現象を,定式化しようとするととたんに難しくなる。ところが,BMは二つの倒立効果という切り口から,少なくとも二つの処理過程が存在することが示唆される。すなわち,動きから知覚される形態処理と局所,特に足の動き情報に関する二つの倒立効果から処理特性を明らかにすることができる。以降,本稿ではヒトの関節に光点を装着した歩行運動としてBMを定義する。
一つ目のBM倒立効果は,顔知覚に見られる倒立効果−正立顔を180度ひっくり返すことにより,顔の知覚が阻害されるという効果である(Sumi, 1984)。すなわち,正立BMを180度回転することで,それはもはや歩行している「ヒト」はなく,ある報告者によれば,竜巻のような点の動きにしか見えないというのである。
もう一つの倒立効果は,形態とは独立の,足の動きに基づく倒立効果である(Westhoff & Troje, 2006)。これは,BMの各光点を空間的にランダムに配置することによりヒトの形状を検出できなくても,左右どちらに進行しているかを判定することができる。特に,足の動きの軌跡が,進行方向を判断する際の手がかりとなることが知られている。
このような足の動きの軌跡を倒立させると,進行方向弁別課題成績はチャンスレベルへと落ちてしまう。つまり,足の動きそれ自体に進行方向に関する情報が埋め込まれているというわけである。このような現象が観察された理由として,我々の足の動きは筋骨格系の制約と重力によって生み出されていることが考えられる。つまり,ヒトが歩く際にはこの二つの制約によって生成される特徴的な動きがあり,我々の視覚系はそのような特徴的な情報を抽出可能であることを示唆する。
著者らはさらに研究を推し進め,これら二つの倒立効果は必ずしも独立ではなく,相互作用することを見出している。すなわち,足の動きの空間的な位置によって,動きの倒立効果が変調することを見出している。例えば足の動きの位置が画面の上方に位置するときには,足の動きが正立・倒立にかかわらず,進行方向の正答率がチャンスレベルとなるが,足の動きの位置が画面の下方に位置するときには,足の動きが正立の場合は正答率が9割近くなる一方で,倒立の場合には正答率は2割近くなる。すなわち,足の動きの空間的な位置によって倒立効果が変調される(Hirai et al., 2011)。
では,このようなBMを我々はいつから区別できるのだろうか? 発達の視点からBM知覚に最初に挑んだのがFoxとMcDaniel(1982)である。Science誌に掲載されたその論文では,2, 4, 6ヵ月児を対象とし選好注視法により検討し,結果,4ヵ月児で既に正立のBMへの選好があることを明らかにした。さらにその後,視覚経験を有しないと考えられる初生雛や新生児を対象とした論文が発表された。一つ目は暗闇で初生雛にメンドリBMとランダムな光点運動を同時に提示した場合,メンドリBMへの選好が見られた(Vallortigara et al., 2005)。ところが,BMとBMの各光点を空間的にランダマイズした光点の映像を呈示した場合には,BMへの有意な選好は見られなかった。すなわち,初生雛は局所的な生物らしい動きを手がかりとしていることが考えられる。さらにヒト新生児でも初生雛と同様に,倒立したメンドリのBMやランダムな光点運動よりも正立のメンドリのBMへの選好が引き出されることが報告されている(Simion et al., 2008)。
著者らは,乳児・児童を対象とした研究を進め,BM処理の発達変化を追跡した。8ヵ月児を対象にBMを知覚している際の脳活動を計測した研究では,正立BMに対する神経活動が統制刺激よりも有意に増大することを見出している(Hirai & Hiraki, 2005)。さらにこのような生後初期に見られるBM知覚への鋭敏性はその後も発達変化し続けることを見出している。著者らは,7〜14歳を対象とした脳波研究において,10歳あたりまで脳活動が変化し続けることを見出している(Hirai et al., 2009)。このような脳活動の発達変化は顔知覚処理においても知られており,BM知覚処理も顔知覚処理と同様の発達過程を辿る可能性が考えられる。これらをまとめると,BMの処理は視覚経験のない新生児においてもみられるが,その知覚処理は成人に至るまで変化し続ける可能性がある。
このようなBM知覚発達過程に関して,実験に基づくエビデンスが蓄積されているものの,背後にある発達の原理については不明であった。そこで,著者らは近年BM知覚処理発達に関する理論モデルを提案した(Hirai & Senju, 2020)。この発達処理モデルは,MortonとJohnsonが1991年に提案した,顔認知発達に関する二段階処理モデルに基づいている。このモデルでは,CONSPECとCONLEARNという二つのモジュールを想定している。CONSPECは目と口の配置に基づき顔への空間的な定位を担い,CONLEARNではCONSPECにより入力された顔知覚処理の学習に関するモジュールである。著者らはこのような2プロセスモデルを参考に,二つのモジュールからなるBM知覚発達モデルを構築した。一つはStep Detectorと呼ばれる,大まかな歩行情報を処理するモジュールであり,これは主として皮質下が関与することを想定している。もう一つはBodily Action Evaluatorと呼ばれる,ヒトの動作・行為を詳細に処理するモジュールであり,主として大脳皮質が関与することを想定している(図2)。
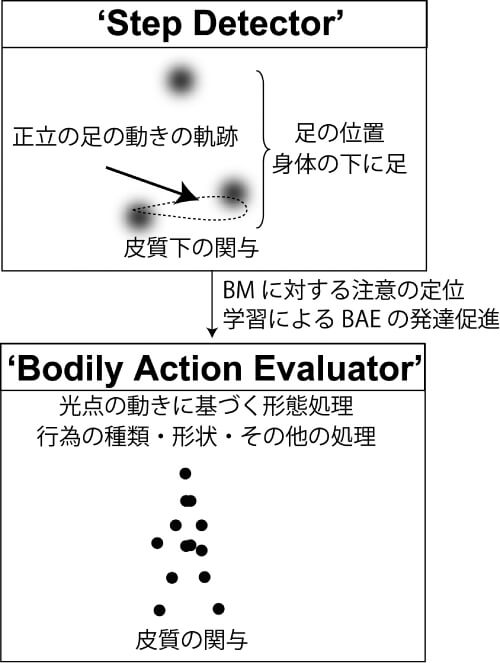
Step Detectorは皮質下にあるゆえ生後すぐに機能し,主としてヒトの歩行運動に関する抽象的な特徴量を検出する機構を想定している。具体的には,正立の足の動きと足の空間的な位置の組み合わせがこのモジュールを駆動する。この二つの特徴量が入力された場合に,その対象へ注意を向ける機構である。一方,Bodily Action Evaluatorでは,行為の詳細な特性を処理するモジュールである。このモジュールはStep Detectorにより注意が向けられることにより,歩行運動への入力が増える。これにより,光点運動から抽出される高次の特徴量を主として処理する。このような二つのモジュールを想定することにより,これまで報告されているBM知覚処理に関する知見を矛盾なく説明することが可能となった。
冒頭でも述べたように,BMの知覚処理により社会脳の一部が賦活されることが知られている。このような経緯から,BMは上述の定型発達児を対象とした研究だけでなく,非定型発達児を対象とした研究も多数行われている。自閉スペクトラム症の診断がつく前の2歳児において,正立BMと倒立BMを提示した場合,定型発達児では,正立のBMへの選好が見られたものの,自閉スペクトラム症児では正立・倒立への選好が見られなかったことが報告されている(Klin et al., 2009)。著者らは定型発達児ならびに自閉スペクトラム症児を対象とした脳波計測実験を進めた(Hirai et al., 2014)。結果,BM知覚処理に関連した脳波は,定型発達児では正立BMに対する神経活動が統制刺激よりも増大するのに対し,自閉スペクトラム症児ではそのような違いが見られなかった。したがって,自閉スペクトラム症児では,光点の動きから形態情報を抽出する処理に不得手さがあるのかもしれない。しかしながら,現時点において何故このような非定型な処理となるかについては十分解明されておらず,さらなるエビデンスの蓄積だけでなく,原理についても明らかにしていく必要がある。
著者らは上述のようなBM知覚発達メカニズムだけでなく,他者の動きが乳児の学習に果たす役割についても検討を始めている(Hirai & Kanakogi, 2019)。これまで,乳児へのアイコンタクトや乳児に向けた発話などの社会的手がかりが重要であるとの研究が蓄積されているが,本当にそれだけだろうか? 実際,養育者が乳児に向けて例えば物体の使い方などを教示するときには,養育者の動作が誇張されたり,繰り返されたりする動作であるモーショニーズが見られる(Brand et al., 2002)。また逆に,乳児もモーショニーズを選好することが報告されている。
しかしながら,モーショニーズのような誇張された動きへの選好は視覚的な顕著性が高いがゆえに選好したとも考えられる。これを踏まえ著者らは,顕著性をできるだけ揃えたコミュニカティブな動きで,乳児の選好や学習を変調するかについて調べた。日常生活のコミュニケーション様式に着想を得て,手の振り方向の影響について検討した。4ヵ月児を対象に乳児に働きかけるような,手を水平方向に動かす映像を統制条件として垂直方向に動かす映像を対提示した結果,水平方向への選好が見られた。さらに9ヵ月児では水平方向の腕振りが乳児の物体学習を促進することを見出している。
このように,我々は生物らしい動き,歩行情報へ注意を向ける機構を生まれながらに兼ね備えており,生後の発達により他者の動作に関する処理が精緻化し,さらにはそのような動き情報を利用することにより効率良い学習を可能としているのではないかと著者は現時点で考えている。しかしながら,顔や視線などの社会的刺激に関する研究に比べ,ヒトの動き情報に関する研究はまだ十分とは言い難い。今後,実験に基づくエビデンスの蓄積とともに理論整備が重要であると考えられる。
近年では,深層学習などの研究が急速に発展し,モーションキャプチャーといった大掛かりな装置は不要となり,ビデオカメラで計測された映像からリアルタイムで身体動作情報を抽出できるまでになっている。さらには,このようにして得られた大量のデータからヒト身体動作の様々な特徴量を精度良く抽出できることが期待される。これにより,ヒトの動きに関する新たな研究の展開が可能となると考える。
ヒトの動きに関する研究領域は上述の心理学だけにとどまらず,神経科学,コンピューターサイエンス,医学,アート,映画やゲームなどのエンターテインメント領域などにまたがる学際的な学問分野である。今後,多くの研究者がヒトの動きに関する研究領域に参画することにより研究が大きく進展することが期待される。
文献
- Brand et al. (2002). Dev Sci, 5 , 72-83.
- Fox & McDaniel (1982). Science, 218 , 486-487.
- Hirai & Hiraki (2005). Brain Res Cogn Brain Res, 22 , 301-304.
- Hirai et al. (2009). Neuroscience, 161 , 311-325.
- Hirai et al. (2011). Psychol Sci, 22 , 1543-1549.
- Hirai et al. (2014). Res. Autism Spectr Disord, 8 , 1623-1634.
- Hirai & Kanakogi (2019). Dev Sci, 22 , e12787.
- Hirai & Senju (2020). Neurosci Biobehav Rev, 111 , 114-124.
- Johansson (1973). Percept Psychophys, 14 , 201-211.
- Klin et al. (2009). Nature, 459 , 257-261.
- Morton & Johnson (1991). Psychol Rev, 98 , 164-181.
- Simion et al. (2008). Proc Natl Acad Sci U S A, 105 , 809-813.
- Sumi (1984). Perception, 13 , 283-286.
- Vallortigara et al. (2005). PLoS Biol, 3 , e208.
- Westhoff & Troje (2006). Curr Biol, 16 , 821-824.