
刊行物





- HOME
- 刊行物のご案内
- 心理学ワールド
- 104号 空間認知の科学 最前線
- 空間認知の神経メカニズム
【特集】
空間認知の神経メカニズム

佐藤 暢哉(さとう のぶや)
Profile─佐藤 暢哉
広島大学大学院生物圏科学研究科博士課程修了。博士(学術)。京都大学霊長類研究所共同利用研究員,日本学術振興会特別研究員(日本大学),科学技術振興機構博士研究員,日本学術振興会海外特別研究員(ロチェスター大学)などを経て,2009年関西学院大学文学部総合心理科学科准教授,2014年より現職。専門は行動神経科学。著書に『神経・生理心理学(シリーズ心理学と仕事2)』『なるほど! 心理学実験法』(ともに分担執筆,北大路書房)など。
空間認知の神経メカニズムの研究といえば,場所細胞とグリッド細胞の研究が2014年のノーベル生理学医学賞を受賞したことを記憶されている方もあるだろう。どちらも動物が特定の場所にいるときに反応するもので,細胞が反応する様子がヒートマップ形式で示されることが多い。印象として分かりやすい示しかたのため,その細胞が場所を表象しているということでわかった気になるかもしれないが,少し考えると,必ずしも当然のことではない。本稿では,こうした空間認知の理論的側面について少し議論してみたい。
場所細胞
大脳辺縁系の一部位である海馬のニューロンには,生体がある空間内の特定の領域に位置している際に活発に活動電位を発するものがある[1]。このような細胞は場所細胞(place cells)と呼ばれ,生体が場所フィールドと呼ばれる領域内にいるときに盛んに活動する(図1A)。その際,環境内での頭の向きは関係がなく,どの方向を向いていても場所フィールド内に入ると活動が盛んになる。ほとんどの場所細胞の場所フィールドは,その空間内でたった一か所である。
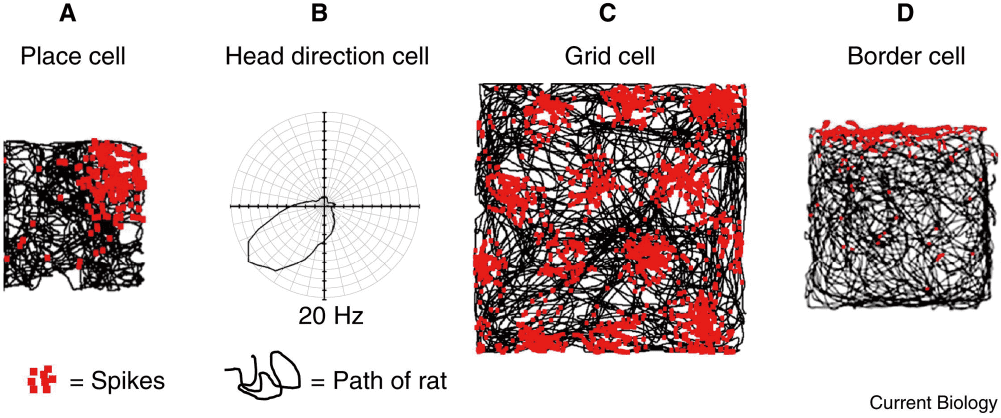
A,C,Dの黒い線はある空間におけるラットの走行軌跡で,赤いドットは,当該の細胞がスパイクを発した(細胞が反応した)場所を示している。Bの黒い線は,ラットがその方向に頭を向けた際の細胞の反応の強さ(スパイクの頻度)を示している。線が中心から外側にあるほど反応が強い。文献3より許諾を得て掲載。D(境界細胞)については本記事では扱っていない。
海馬の場所細胞については様々な特徴が知られており,その一つにリマッピングがある。リマッピング(グローバル・リマッピング)は,場所細胞が反応する場所フィールドが,外的環境の変化に応じて再配置される現象である[2]。同じ実験室内であっても,異なる形状の実験箱に入れられると,細胞が反応する実験室内の相対位置が変化する。反応する場所が変わるので,位置の表象としては不安定な情報ともいえるが,一定数の細胞ポピュレーション全体としては,空間あるいは文脈に依存した当該空間内の位置を表象していると考えられている。
グリッド細胞
海馬への主要な情報の入力源は嗅内皮質である。齧歯類の嗅内皮質は内側部と外側部に分けることができるが,空間的な情報は主に内側部が扱っている。嗅内皮質内側部には,反応する領域が場所細胞のように一か所ではなく,複数の反応領域が正三角形状パターンに整然と並ぶグリッド細胞(grid cells)が存在する[4](図1C)。反応領域のパターンは違うが,いくつかの点でグリッド細胞は海馬の場所細胞と類似している。たとえば,海馬場所細胞と同様に,複数のグリッド細胞の活動から,その活動を記録している動物の現在地をデコードするのに十分な情報を得ることができる。また,各グリッドの各反応領域の大きさ,およびグリッドの間隔が嗅内皮質内側部の背側から腹側にかけて増大することも,海馬の場所細胞の場所フィールドが背側部では小さく,腹側部では大きいことと一致する。近くにあるグリッド細胞が反応する領域は少しずつずれているが,そのずれの方向には法則性がないことも,隣り合う場所細胞の場所フィールドの位置に法則性が認められないことと類似している。
頭部方向細胞
場所細胞やグリッド細胞の活動は空間内での位置を表象しており,空間に対する身体の向きとは関係ない。つまり,どこにいるのかという情報は持っているが,どちらを向いているのかという情報は持っていない。これに対して,Papez回路に含まれる後海馬台,乳頭体,視床前核などにみられる頭部方向細胞は環境内での頭の向きによって応答性を変化させ,環境内での場所には依存しない[5]。つまり,その細胞の好みの方向を向いていれば,どこにいるかは関係なく反応する(図1B)。このように,頭部方向細胞と場所細胞とは表裏の関係にある。典型的な頭部方向細胞は単一の好みの方向に強い選択性を示す。
細胞応答特性のモデル
頭部方向細胞の選択性の形成過程については,連続アトラクターネットワーク(continuous attractor network: CAN)を基盤とした理論が提唱されている[6]。アトラクターネットワークはニューラルネットワークの一つで,複数の安定状態が存在し,どの安定状態に落ち着くかは,その時々のネットワークの活動パターンに依存する。個々の神経細胞ユニット同士が隣接したユニットと相互に局所的な興奮性の結合を有し,かつ全体的には過活動を抑制するようなフィードバック結合があるニューラルネットワークがあるとする。このようなネットワークは,一つの隆起(bump,あるいはactivity packetと呼ばれる)を持つポピュレーション活動として安定することが知られている(図2上)。このネットワーク内の特定の細胞集団のみが活性化している安定状態(隆起)は自然発生的に生じるもので,このままで何かの情報を表象しているわけではない。
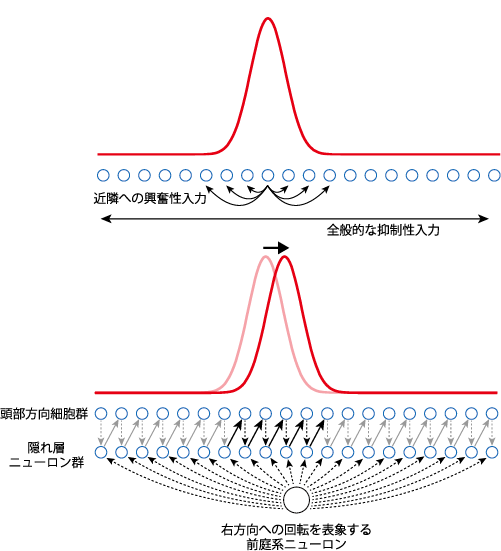
上:近隣のニューロンへ勾配をつけた興奮性の入力,および過活動を抑えるための全般的な抑制性の入力が想定されている。このようなネットワークは一つのbumpを示す状態で安定する。下:頭部方向細胞の連続アトラクターネットワークモデル。隠れ層のニューロン群には,頭部方向細胞群からの現在情報の入力と,前庭系からの頭部回転の情報が入力される。隠れ層ニューロン群の出力は頭部方向細胞群へシフトする形でなされる。このようにして,頭部方向の状態情報が更新される。この図では右方向への頭部方向の変化のみが示されているが,左方向の変化については別の隠れ層が想定される。
ここで,このネットワークに中間層(隠れ層)を加える。隠れ層のユニットは,元のネットワークを構成する各ユニットからの入力に加えて,なんらかのパラメータ(頭部方向細胞の場合は頭の向き)が変化したという情報を受ける。そして,その隠れ層の出力によって,元のネットワーク状態が更新されるような結合があると(図では,右方向へユニット一つ分シフトした形での出力結合),そのネットワークは,そのパラメータの現在状態を表象するものとなる(図2下)。ネットワークが,標的とするパラメータの現在の状態を表象するようになるのは,そのパラメータの変化についての情報が与えられ,そして,その変化の情報によって状態が更新されるからである。頭部方向細胞のCANモデルでは,隠れ層のユニットは前庭系からの頭部方向の変化情報の入力と,頭部方向細胞から現在の頭部方向の情報を受け取る(頭部方向細胞群から隠れ層への入力は,各ユニットから出る下向き点線の矢印で示している)。そして,隠れ層から頭部方向細胞層へ出力が戻されるわけだが,この出力は変化に応じてシフトする形で行われる(図の例では一つ右隣のユニットに出力が与えられる)。このことで,頭部方向細胞群の状態が更新され,隆起が少しシフトした安定状態へと移行する。
場所細胞やグリッド細胞のように空間内の特定の場所に対する反応する特性も,CANを2次元に拡張することで形成できる。この場合,ネットワーク状態を更新するために,2次元平面上での移動方向の情報と,もとの場所細胞群から入力される現在の位置状態とを集約する隠れ層が必要となってくる。実際に,グリッド細胞の一部には,グリッド状に並んだ位置への反応が頭部方向によって変調されるような特性を示すものがあり[7],こうした細胞は隠れ層ユニットに相当する可能性がある。
頭部方向細胞は,1次元のCANによって構成できるが,この際,方向という周期性のある性質と一致して,細胞ユニットはリング状に構成されていると考えられている(図3上)。2次元平面の場合も,無限に広がる平面を表象することは現実的ではないため,1次元の場合と同様に,それぞれがある程度の広さの反応領域を持つ細胞ユニットの境界を反対側の境界とつなげるとよい。その結果,細胞の反応の構成はトーラス状になる(図3下)。このトーラス表面上に一つの場所フィールドがあるとすると,実空間上では,反応する場所フィールドが一定の間隔で繰り返される形になると予測される[6]。この予測と一致した反応様式を示すのがグリッド細胞である。つまり,グリッド細胞の反応する場所フィールドが規則正しく配置されていることは,それが2次元的なCANによって構成されているためだといえる。実際に,内側嗅内皮質のグリッド細胞の反応様式がトーラス状の構造を持つことも報告されている[8]。
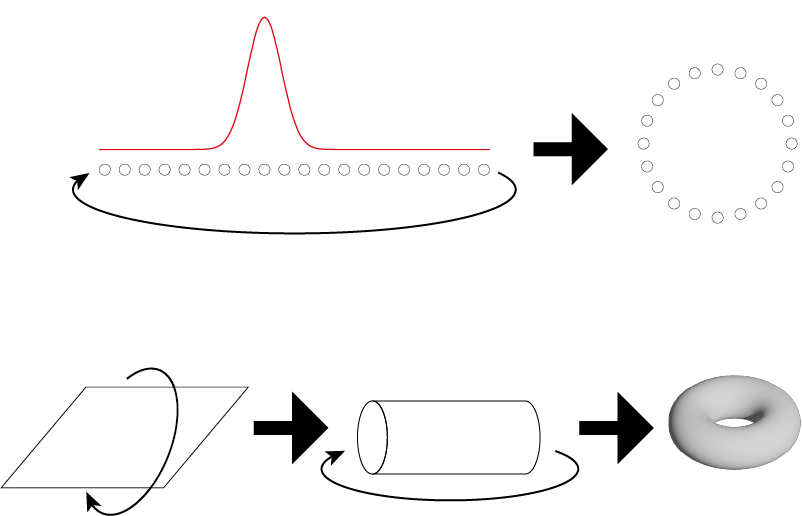
上:頭部方向細胞では,リング状にユニットが配置される。下:二次元平面の場合,境界をなくすためには両軸のエッジを結合すればよい。図中平面の縦方向の両端を結合すると円筒形になり,さらに円筒の両端を結合すればトーラス状になる。
グリッド細胞には,ある一定の広がりを持った領域が表象されているが,生体がその領域を出ると同様の反応領域が繰り返し表象されることになる。そのため,同様に繰り返し出現する反応領域のうち,どれが自分のいるものなのかが区別できなくなってしまう。しかし,個々のグリッド細胞が集まったポピュレーションとしては移動についての情報は持っているので,2次元平面上での生体の移動が表現され得る。こうした情報を利用することによって経路統合(path integration)が実現可能である。
経路統合とは,自身の移動の情報(どの方向にどの程度の距離移動したのかという情報の系列)を統合することで,出発地点との相対位置として現在地点を同定することである。空間内での位置の情報は,GPSのように絶対的な位置情報を持つということは考えにくい。巣を持つ動物の多くは,主に巣を基準点として,移動情報を統合することで,巣から出て一定の移動後の現在位置を同定すると考えられる。
経路統合では,現在地情報を更新していくために,自身の運動の情報を随時蓄え,それを統合する計算をし続ける必要がある。常に情報が更新され続けなければならないため,誤差も加算的に増大していくことになる。そのため,長距離・長時間の移動後には,経路統合によって算出された位置は,実際の空間位置とずれてしまうことになる。これを避けるために,ランドマーク(空間内の特定の位置に紐づいた外部情報)を用いた現在地情報の校正がなされると考えられている。頭部方向細胞では,このような校正プロセスに当たる活動がみられることも知られている[5]。
海馬の場所細胞について
グリッド細胞と同様に,海馬の場所細胞の場所フィールドもCANから構成されることが提唱されているが[9],グリッド細胞に比べると説明できないことも多いと考えられているようである。リマッピング現象はCANの安定状態の遷移に相当するのかもしれないが,実際にどのように遷移が生じるかなどの詳細はまだ明らかとはいえない。典型的な場所細胞では,当該環境内に一つの場所フィールドがみられ,グリッド細胞のような繰り返しパターンはみられない。つまり,一つの場所細胞が,空間内に複数の反応領域を持つことはない。実空間において,ある環境を表象する際,境界もなく無限に表象空間が広がっているということは考えにくく,我々が実際に実空間の表象を形成する際は,ある境界によって当該空間を切り取っていると思われる。海馬の場所細胞は,このように何らかの形で区切られた空間の表象を構成していると思われる。
この区切られた小空間は「文脈」に相当するのかもしれない。つまり,場所細胞は現在位置というよりも,当該空間を文脈として表象することを行っている可能性がある。この際,場所細胞ポピュレーションに含まれる個々の細胞の場所フィールドが安定化することが当該空間の文脈表象の完成に当たると思われる。海馬と文脈表象との関わりは,これまでも多くの脳損傷実験などで示されてきたことでもある。
グリッド細胞群が経路統合によって現在位置を表象しているとすると,別の形式であるとはいえ,海馬において現在位置情報をもう一度形成する意味はあまりないように思える。実際に,海馬の場所情報には現在位置以上のなにかがあると考えられており,その一つがエピソード記憶への関与である。それを裏づける現象の一つに,かつて通った(あるいはこれから通る)経路に沿った場所フィールドが系列的に,かつ瞬時に再活性化されるリプレイ現象がある[10]。
場所細胞様活動の種による違い
場所細胞およびグリッド細胞はラットにおいて発見されたこともあり,その研究のほとんどが齧歯類を対象としたもので,霊長類における報告は多くはない。サルの海馬では,場所細胞よりも景観細胞(spatial view cell)と呼ばれるニューロン活動が優勢だといわれている[11]。これは,空間内の場所ではなく,視線を向けた先にある景色に対して反応する細胞である。この景観細胞の反応様式は,場所細胞(空間内の特定の場所にいるときに反応)や頭部方向細胞(特定の方向に頭を向けているとき反応)とも異なっている。なぜサルの海馬では場所細胞ではなく景観細胞が優勢なのかは未だ不明だが,齧歯類が外界情報の取得に嗅覚系や触覚系に頼る割合が高く,主に経路統合によって現在地を同定するのに対して,霊長類では視覚系に重きを置き,眼球運動系が発達していることから,ランドマークやシーンの視覚的認知を通した現在地の同定が主であることと関係するのかもしれない。
空間情報は元来感覚情報に根差したものではない。時間と同様に,空間情報を直接検出する感覚器官は存在せず,空間情報の表象は感覚情報を利用して内的に作り出す必要がある。そのため,感覚情報やその処理系の重みづけの違いが空間情報処理系に反映されることは当然なのかもしれない。このことは,神経細胞の反応様式の違いだけではなく,現在地の同定やナビゲーション方略にも影響すると予測され,実際にそれらの種差を示唆する報告もなされている。空間認知の神経メカニズムについては齧歯類における研究が華々しいためか,こうした点はあまり着目されない印象もあるが,人間の空間認知を考える上では重要であろう。
- 1.O’Keefe, J., & Dostrovsky, J. (1971) Brain Res, 34, 171–175.
- 2.Muller, R. U., & Kubie, J. L. (1987) J Neurosci, 7, 1951–1968.
- 3.Marozzi, E., & Jeffery, K. J. (2012) Curr Biol, 22, R939–942.
- 4.Hafting, T. et al. (2005) Nature, 436, 801–806.
- 5.Taube, J. S. (2007) Ann Rev Neurosci, 30, 181–207.
- 6.McNaughton, B. L. et al. (2006) Nature Rev Neurosci, 7, 663–678.
- 7.Sargolini, F. et al. (2006) Science, 312, 758–762.
- 8.Gardner, R. J. et al. (2022) Nature, 602(7895), 123–128.
- 9.Samsonovich, A., & McNaughton, B. L. (1997) J Neurosci, 17, 5900–5920.
- 10.Wilson, M. A., & McNaughton, B. L. (1994) Science, 265, 676–79.
- 11.Rolls, E. T. et al. (1997) Eur J Neurosci, 9, 1789–1794.
- *COI:本記事に関連して開示すべき利益相反はない。
PDFをダウンロード
1